luni, 28 decembrie 2015
marți, 15 decembrie 2015
marți, 10 noiembrie 2015
marți, 20 octombrie 2015
PERSPECTIVA UTILIZĂRII VIRUSULUI POLIEDROZEI NUCLEARE ÎN COMBATEREA OMIZII PĂROASE A STEJARULUI
PERSPECTIVA UTILIZĂRII VIRUSULUI POLIEDROZEI NUCLEARE ÎN COMBATEREA OMIZII PĂROASE A STEJARULUI
Pascaru Alexandru;
Lungu Andrei; Voloșciuc Leonid
Institutul de Genetică, Fiziologie şi Protecţia
Plantelor
Abstract
In Republic of
Moldova Lymantria dispar L. provokes massive defoliation in forests of
oaks and other forest species, and in fruit orchards. The use of pesticides to
combat, provokes serious problems for
environment, but and human health. As an alternative method to combat,
was proposed using baculoviral preparations, that ensuring a high level of
effectiveness, showing properties of specificity. In many countries with
advanced forestry services also applied to surfaces prepared imposing. In the
Republic of Moldova were carried out investigation on highlight biological
agents and demonstrate the use of preparations baculovirale, but level the
application of these means now remains reduced, recording various problems, who
awaiting approach and resolve [1, 9, 12].
Keywords: baculovirus,
biologic control, ecology, forest ecosistems, insecticides,
Lymantria dispar.
Introducere
În pădurile de foioase din ţara noastră şi din alte ţări, se
dezvoltă un număr mare de dăunători silvici. Unul dintre aceşti defoliatori
este omida-păroasă-a-stejarului, larva speciei Lymantria dispar L. În urma atacurilor ai acestei
specii, arborii îşi încetinesc creşterea, iar vitalitatea lor, ca consecinţă
poate scădea treptat, favorizîndu-se în acest mod dezvoltarea agenţilor fitopatogeni
secundari [6, 8, 9,].
Larvele de Lmantria dispar, acestuia sunt capabile să distrugă periodic
(odată la 8-10 ani), sectoare forestire de zeci de mii de hectare silvice.
Ţinând cont că cvercineele predomină în ecosistemele silvice din Republica
Moldova, ocupând 39,6% din fondul forestier, impactul dăunătorului poate devine
considerabil [1, 3, 5].
Protejarea
integrată a pădurilor include un complex de măsuri (silviculturale, biologice,
chimice), crearea de culturi forestiere rezistente la atacul dăunătorilor şi
aplicarea de pesticide şi produse biologice în scopul prevenirii vătămărilor şi
diminuării populaţiilor de insecte dăunătoare [14, 15].
Este
necesar de menţionat că pesticidele, fiind ingrediente
străine ale ecosistemelor naturale, cauzează dereglarea relaţiilor biocenotice,
întrerupând lanţurile trofice interspecifice, reduc densitatea populaţiilor
entomofaunei utile. Altă urmare negativă este apariţia liniilor de insecte
dăunătoare, rezistente faţă de unul sau chiar mai multe pesticide [5, 11, 18].
Astfel drept metodă alternativă de
combatere a dăunătorilor silvici poate fi utilizarea Virusului Poliedrozei
Nucleare, manifestând proprietăţi de
specificitate, în special pentru Lymantria dispar [10].
Virusul Poliedrozei
Nucleare (VPN), face parte din familia baculovirusurilor, genul
alfabaculovirusurilor, specia Borrelinavirus euproctis [4]. Această specie provocă poliedroza nucleară a
omizilor la specia L. dispar [16].
Materiale și metode
Cercetările se efectutiază pe larvele Lymantria
dispar de vârsta a II-a şi a III-a. După ziua a treia de la infectare se începe evidenţa mortalităţii
larvelor pe variante. Determinarea
larvelor bolnave se efectuiază după simptomele respective [2, 5].
Determinarea titrului soluţiilor virotice se
efectuiază cu ajutorul camerei Goreav sau în preparatele fixate şi colorate, la
microscopul optic cu cameră. Determinarea titrului VPN se efectuiază după formula
(îndrumări metodice, Ciuhrii M. G. 1991):
Σ SPVC în 100 pătrate mici
x 4 000 000
T= -------------------------------------------------- *
K sau T= 105 x a x b
100
Unde: T- titrul
virusurilor; K- gradul diluării suspensiei virale; a- suma SPVC în 40 de
pătrate mici; b- nivelul diluţiei suspensiei virale.
Testarea în condiţii de laborator şi în
câmpul de experienţă s-a efectuat în 3 variante, fiecare câte 4 repetiţii, în
conformitate cu cerinţele generale de acest gen [11, 13].
Eficienţa biologică a suspensiilor virale
se determină după formula Abbot, care prevede mortalitatea naturală a
insectelor:
Me-Mc
Ab = --------------------- * 100
100-Mc
Unde: Ab- indicatorul mortalităţii;
Me-
numărul de larve moarte în experienţă;
Mc-
numărul de larve în control.
Omizile, după eclozare din ouă, se hrăneşte
timp de 2 săptămâni, până ating vârsta II-a, apoi se separă în variante
rendomizate câte 40 de larve, se plasează în borcane de sticlă sau containere
mici de plastic. La această etapă începe experienţa propriu-zisă. Evidenţa
mortalităţii larvelor Lymantria dispar se efectuiază până la a 15-a zi
[2, 7, 9].
Rezultate și discuții
La nivel mondial împotriva acestui dăunător sunt aplicate
insecticide virotice, spre exemplu în SUA- preparatul Djipcec şi Virex, în
Canada- preparatul Lecontvirus, etc. Un şir de preparate analoage sunt
utilizate în Europa şi Asia [17, 18]. Iar în Republica
Moldova au fost efectuate cercetări asupra evidenţierii agenţilor biologici şi
demonstrării perspectivelor utilizării preparatelor baculovirale, dar
nivelul aplicării acestor mijloace deocamdată rămâne unul redus, înregistând diverse
probleme, care-şi aşteaptă abordarea şi suluţionarea [10].
Creşterile
pe hrană naturală permit obţinerea de material biologic numai în cursul
sezonului de vegetaţie.
În
anul 2014 pontele au fost izolate de pe teritoriul Ocolului Silvic Străşeni şi
Dendrariul din Chişinău. În rezultatul experienţei a fost obţinută o nouă
cantitate de biomasă virală, care urmează a fi folosită în următoarele experienţe. Rezultatele cercetărilor sunt prezentate în
tabelul nr.1 de mai jos.
Tabelul nr. 1
Rezultatele obţinute în baza cercetărilor anul 2014
Variante şi repetiţiile lor
|
Titrul suspensiei SPVC/ml
|
Numărul
de larve moarte
|
Procentul
mortalităţii
|
Indicele Abbot
|
Varianta I
|
||||
Repetiţia 1.1.
|
T= 105 x
467 x 101
|
39
|
97,50%
|
97,05%
|
Repetiţia 1.2.
|
T= 105 x
376 x 101
|
39
|
97,50%
|
97,05%
|
Repetiţia 1.3.
|
T= 105 x
432 x 101
|
39
|
97,50%
|
97,05%
|
Repetiţia 1.4.
|
T= 105 x
392 x 101
|
40
|
100%
|
100%
|
Martor
|
-
|
6
|
15%
|
-
|
Varianta II
|
||||
Repetiţia 2.1.
|
T= 105 x
38 x 102
|
39
|
97,50%
|
97,22%
|
Repetiţia 2.2.
|
T= 105 x
44 x 102
|
40
|
100%
|
100%
|
Repetiţia 2.3.
|
T= 105 x
43 x 102
|
38
|
95%
|
94,44%
|
Repetiţia 2.4.
|
T= 105 x
66 x 102
|
40
|
100%
|
100%
|
Martor
|
-
|
4
|
10%
|
-
|
Varianta III
|
||||
Repetiţia 3.1.
|
T= 105 x
14 x 103
|
38
|
95%
|
93,94%
|
Repetiţia 3.2.
|
T= 105 x
6 x 103
|
36
|
90%
|
88,57%
|
Repetiţia 3.3.
|
T= 105 x
5 x 103
|
39
|
97,50%
|
97,14%
|
Repetiţia 3.4.
|
T= 105 x
5 x 103
|
38
|
95%
|
93,94%
|
Martor
|
-
|
5
|
12,50%
|
-
|
Concluzii
1. O sursă de poluare a
ecosistemelor silvice reprezintă insecticidele chimice. Această poluare prezintă pericol
pentru speciile rare de plante şi animale. Astfel poate apărea un dezechilibru în ecosistemele
silvice.
2. În dependenţă de titrul suspensiei
virale, care a fost utilizat, la începutul experienţei se evidenţiază o diferenţă
procentuală a mortalităţii larvelor, ca la sfîrşitul experienţei, mortalitatea
în variante şi repetiţii devine asemănătoare, astfel pentru infectarea în masă a
omizilor este necesară o concentraţie minimală.
3. Majoritatea omizilor mor în
primele zile ale experienţei.
4. Aplicarea pe scară largă a
insecticidelor virotice poate deveni o realitate doar la organizarea unui
sistem larg de laboratoare biologice moderne, destinate producerii lor;
Bibliografie
1. CIUHRII, Mircea. Utilizarea baculovirusurilor în controlul
densităţii populaţiilor de insect nedorite. ISSN 2068-4096. Volumul 1,
Nr. 1 . Lucrările Sesiunii ştiinţifice de la Neptun-Comorova, 2010. P. 241-245.
2. ДОСПЕХОВ Б. 1989. Методика полевого опыта. М.: Колос. 416с.
- GIURGIU, V.
(2011). "Revista pădurilor
(Journal of forests) 125 years of existence". Rev. pădur. 126 (6): 3–7. ISSN 1583-7890.
- LIŢESCU M. Starea de sănătate şi principalii
dăunători ai pădurilor din raza Ocolului Silvic Costeşti (păduri de foioase). Revista ,,ECOS” 20/2008, P.
124-128.
- NICOLAESCU O., VOLOŞCIUC L. Rolul preparatelor
virale in menţinerea densităţii populaţiilor de Lymantria dispar L.
Lucrări ştiinţifice UASM Volumul 24 (2). 2010. P. 471-476.
6.
OLENICI N. (2009). Silvicultura
durabila intr-un mediu in schimbare. Bucureşti. Tehnica Silvica. 240 P. ISBN: 9789738893894.
7.
POGUE, M., SCHAEFER,
P. A Review of Selected Species of Lymantria Hüber [1819] (Lepidoptera: Noctuidae:
Lymantriiae) from Subtropical and Temperate Regions of Asia, Including the
Descriptions of Three New Species, Some Potentially Invasive to North America.
Washington, 2007. 232 p.
8. REARDON, R., PODGWAITE, J. P.
and R. T. ZERILLO.1996. GYPCHEK - The gypsy moth nucleopolyhedrosis virus
product. USDA Forest Service publication FHTET-96-16. 420 p.
9.
SIMIONESCU A., MIHALACHE Gh. (2000). Protecţia pădurilor.
Regia naţională a pădurilor.Ed.Muşatinii. 880p. ISBN 973-99309-7-2.
10.
STATE of the World’s Forests. (2012). Food and
agriculture organization oF the united nations. Rome. 60p.
11.
VOLOŞCIUC L., NICOLAESCU O. Specificitatea baculovirusurilor asupra larvelor de Lymantria dispar L.
Aspecte ştiinţifico-practice a dezvoltării durabile a
sectorului forestier din Republica Moldova. Chişinău. 2006. P.156-160.
12.
VOLOŞCIUC Ë.T. Biotehnologia producerii şi aplicării preparatelor
baculovirale în agricultura ecologică. Chişinău: Mediul
ambiant, 2009, 262 p.
- VOLOŞCIUC L.T. Problemele
identificării şi ameliorării baculovirusurilor. Buletinul AŞM. Ştiinţele
vieţii. 2010. 1(310), 96-106.
14. VOLOŞCIUC, Leonid. Biotehnologia Producerii şi Aplicării Preparatelor
Baculovirale în Agricultura Ecologică. Chişinău, 2009. 234 p.
15. VOLOŞCIUC L.T. Contribuţia ştiinţei şi inovării în fortificarea sectorului
forestier. Academos. Nr.2 (29).
2013. P.114-122.
16. КОЛОСОВ, А. Разработка и
испытания вирусных энтомопатогенных препаратов для защиты растений. Автореферат диссертации на соискание ученой степени кандидата биологических наук. Кольцово. 2011. 20 c.
17. MОЗОЛЕВСКАЯ. Е. Лесная энтомология и лесная фитопатология – неразрывность связи и условие эффективной лесозащиты. Болезни и вредители в лесах россии: век ХХI . Материалы всероссийской конференции с международным участием и v ежегодных чтений памяти Kатаева. Екатеринбург, 20-25 сентября 2011 г. C.3-4.
18. http://www.ncbi.nlm.nih.gov/pubmed/16648963
(accesat pe data 28.08.15).
miercuri, 23 septembrie 2015
Despre ferigi (Partea I)

Generalități. Ferigile şi plantele înrudite cu ele fac parte din
Încrengătura Pteridophyta. Aceste plante au prezentat prima oară caracteristici
neîntâlnite la muşchi şi la alge – un strat exterior de celule, impermiabil la
apă (cuticulă) şi ţesut specializat pentru transportul apei şi hranei între
organele care absorb apa (rădăcinile) şi organele care produc hrana (frunzele) [3].
Răspândirea
ferigilor. Până în
prezent se cunosc aproximativ 12.000 de specii, răspândite pe tot globul, cu o
frecvenţă mai mare în pădurile umede şi mlaştini. Chiar şi în regiunea arctică exista peste 500 de specii special
adaptate pentru a înfrunta condiţiile aspre. Aproape la fel de multe specii au
dezvoltat solzi sau peri pentru a putea. Să trăiască în condiţii de deșert [3].
Altitudinea
nu este nici ea o barieră, şi unele ferigi cresc la înălţimi de până la 5.000
m. Deoarece ferigile în general prospera în condiţii de umiditate, nu este
surprinzător că cele mai multe specii se găsesc la tropice.
Mărimea
ferigilor variază mult. Cele din câmpul ferigilor membranoase sunt atât de mici
încât o colonie întreaga poate să crească sub o singură frunza de mărime medie.
Frondele lor filigrane sunt aproape transparente şi necesită o umiditate foarte
mare pentru a supravieţui. La cealaltă extremă sunt cele aproximativ 700 de
ferigi arborescente. În Noua Zeelandă şi regiunile umede din Australia ele pot
creşte la înălţimi de 15 m iar frondele lor ca de palmier pot avea lungimea de
6 m.
Unele ferigi
plutesc pe ape stătătoare. Ferigile zână (specia Azollă). Care formează covoare
pe suprafaţa eleşteielor în timpul verii, de obicei supravieţuiesc peste iarnă.
Alte tipuri, precum Lygodium. au descoperit că prin căţărare le este asigurată
supravieţuirea, şi se pot căţăra în copaci până la înălţimi de 10 m [1].
Reproducerea ferigilor. Feriga nu are seminţe pentru a se reproduce, aşa cum
au majoritatea celorlalte plante. Aceasta se întâmpla pentru că ea nu are
flori, în schimb, ferigile au dezvoltat un ciclu neobişnuit care nu a fost pe
deplin înţeles până la mijlocul secolului al XlX-lea când, după şapte ani de
cercetări “în timpul liber”, un botanist german, Fiedrich Hofmeister, a reuşit
să explice misteriosul proces.
Ferigile
produc spori-particule ca de praf conţinute în capsule numite sporangi.
Sporangii sunt grupaţi pe partea inferioară a frondelor în grămezi numite sori.
Adesea formează modele maro închis atrăgătoare. O singură feriga poate să aibă
mii de sporangi.
Când ajung
la maturitate, capsulele explodează şi împrăştie sporii, care sunt luaţi de cea
mai uşoară adiere de vânt. Dacă un spor aterizează într-un loc umed şi umbros
care favorizează creşterea el poate să germineze. Un lanţ de celule se formează
în doar câteva zile, dar trec cel puţin câteva luni până când se formează o
plantă în formă de inimă, având de obicei diametrul de circa 6 mm. Aceasta nu
este însă o ferigă. Ea se numeşte protal şi este o mică planta hermafrodită,
care reprezintă generaţia sexuată a speciei.
 [5].
[5].
 [5].
[5].
Protalul este fixat în sol cu rizoizi asemănători cu
rădăcinile şi are organele sexuale ascunse pe partea inferioară. Părţile
masculine, numite anteridii conţin sperma la capătul ascuţit al procalului.
Părţile feminine, numite arhegoane, se găsesc la capătul opus. Fecundata poate
avea loc când o peliculă subţire de apă acoperă protalul, permiţând spermei să
înoate.
După fecundaţie, celula ou se divide pentru a produce sporofitul asexuat, sau feriga familiară, care îşi va purta propriii spori. Apoi protalul moare. Dacă fecundaţia nu a avut loc, atunci protalul va trăi până când scopul său va fi îndeplinit. Întregul proces ciudat de producere a unei ferigi minuscule din praf viu durează aproximativ 9 luni [1].
Gametofitul, cea de-a doua generaţie, poartă numele de protal (tal iniţial) şi este întotdeauna haploid, deoarece îşi începe de fiecare dată existenţa pornind de la un spor postmeiotic; generaţia gametofitică se încheie cu formarea gameţilor. Sporii la ferigi sunt germeni asexuaţi, haploizi, formaţi în sporangi. Aceştia din urmă sunt grupaţi în spice sporangifere terminale numite strobili (Equisetum sp. s.a.), în sori, de obicei protejaţi de un înveliş numit induzie, sau în sporocarpi, corpuri sporangifere de sine stătătoare (Salviniaceae). Ferigile cu un singur tip de spori se numesc izospore iar sporii pe care îi formează, de aceeaşi mărime, poartă numele de izospori. Dintr-un izospor se diferenţiază un protal monoic, de obicei verde, pe care se dezvoltă atât gametangii masculine (anteridii) cât şi feminine (arhegoane). Ferigile cu două tipuri de spori se numesc heterospore iar sporii pe care îi formeză, de mărimi diferite, se vor numi heterospori. Cei mai mici dintre ei au fost numiţi microspori iar cei mai mari megaspori sau macrospori. Microsporii vor genera, la germinare, numai protale masculine alcătuite din câteva celule numite şi microprotale. Unele celule ale protalului masculin se transformă în anteridii. În acestea se vor diferenţia gârneţii masculini pluriciliaţi numiţi spermatozoizi (anterozoizi). Megasporii sau macrosporii, mai mari decât precedenţii, vor genera la germinare numai protale feminine sau megaprotale alcătuite din numeroase celule. Pe aceste protale se formează arhegoanele în care se diferenţiază gameţii feminini numiţi oosfere. Prin urmare, izosporii dau naştere la protale monoice iar heterosporii la protale dioice. Reproducerea sexuată. Protalele provenite din diviziuni succesive ale sporilor, au dimensiuni variate (de la câţiva mm până la 5 cm) şi forme diferite (lamelar, foliaceu, tuberculiform, cordiform, filamentos). De cele mai multe ori protalul se aseamănă cu talul algelor verzi filamentoase. Protalul se fixează de substrat prin intermediul rizoizilor de origine epidermică. Indiferent de formă şi mărime protalul se dezvoltă întotdeauna separat sau independent de sporofit, pe suporturi permanent umede. Pe protale se diferenţiază organele de reproducere, arhegoanele şi anteridiile la ferigile izospore sau arhegoane pe megaprotale şi anteridii pe microprotale în cazul ferigilor heterospore. Fecundaţia şi formarea zigotului. Ca la toate plantele cu gameţi masculini mobili, fecundaţia se produce în mediu lichid. Graţie flagelilor, spermatozoizii ajung la arhegoane, pătrund în interiorul lor şi fecundează oosfera. După fuzionarea celor doi gameţi haploizi se formează zigotul, germene sexuat diploid. Din zigot, în urma a numeroase diviziuni, se formează embrionul, iar din acesta o nouă plantă. Mult timp reproducerea sexuată a pteridofiţelor a rămas necunoscută, motiv, pentru care ele au fost denumite şi criptogame (vasculare). Ciclul de dezvoltare la pteridofite. Cele două generaţii, sporofitul şi gametofitul, îşi duc viaţa independent, alternând una cu cealaltă. Generaţia sporofitică dominantă este reprezentată de planta propriu-zisă. Pe aceasta se diferenţiază sporangii în care se formează sporii haploizi. Din germinarea sporilor rezultă gametofitul, numitprotal, pe care se diferenţiază gametangii masculini şi feminini, în urma fecundaţiei dintre gameţii de sex opus se formează zigotul iar din acesta embrionul (sporofitul tânăr) şi apoi sporofitul de lungă durată. Un astfel de ciclu de dezvoltare, în care alternează două generaţii morfologic diferite, independente, cu ploidii diferite, poartă numele de ciclu digenetic, diplo – haplo - fazic, iar plantele respective diplo - haplonte. Ciclul de viaţă la ferigi se desfăşoară diferit la cele izospore (Polypodium, Equisetum, Lycopodium), comparativ cu cele heterospore (Selaginella, Salvinia, Azolla) [4].
După fecundaţie, celula ou se divide pentru a produce sporofitul asexuat, sau feriga familiară, care îşi va purta propriii spori. Apoi protalul moare. Dacă fecundaţia nu a avut loc, atunci protalul va trăi până când scopul său va fi îndeplinit. Întregul proces ciudat de producere a unei ferigi minuscule din praf viu durează aproximativ 9 luni [1].
Gametofitul, cea de-a doua generaţie, poartă numele de protal (tal iniţial) şi este întotdeauna haploid, deoarece îşi începe de fiecare dată existenţa pornind de la un spor postmeiotic; generaţia gametofitică se încheie cu formarea gameţilor. Sporii la ferigi sunt germeni asexuaţi, haploizi, formaţi în sporangi. Aceştia din urmă sunt grupaţi în spice sporangifere terminale numite strobili (Equisetum sp. s.a.), în sori, de obicei protejaţi de un înveliş numit induzie, sau în sporocarpi, corpuri sporangifere de sine stătătoare (Salviniaceae). Ferigile cu un singur tip de spori se numesc izospore iar sporii pe care îi formează, de aceeaşi mărime, poartă numele de izospori. Dintr-un izospor se diferenţiază un protal monoic, de obicei verde, pe care se dezvoltă atât gametangii masculine (anteridii) cât şi feminine (arhegoane). Ferigile cu două tipuri de spori se numesc heterospore iar sporii pe care îi formeză, de mărimi diferite, se vor numi heterospori. Cei mai mici dintre ei au fost numiţi microspori iar cei mai mari megaspori sau macrospori. Microsporii vor genera, la germinare, numai protale masculine alcătuite din câteva celule numite şi microprotale. Unele celule ale protalului masculin se transformă în anteridii. În acestea se vor diferenţia gârneţii masculini pluriciliaţi numiţi spermatozoizi (anterozoizi). Megasporii sau macrosporii, mai mari decât precedenţii, vor genera la germinare numai protale feminine sau megaprotale alcătuite din numeroase celule. Pe aceste protale se formează arhegoanele în care se diferenţiază gameţii feminini numiţi oosfere. Prin urmare, izosporii dau naştere la protale monoice iar heterosporii la protale dioice. Reproducerea sexuată. Protalele provenite din diviziuni succesive ale sporilor, au dimensiuni variate (de la câţiva mm până la 5 cm) şi forme diferite (lamelar, foliaceu, tuberculiform, cordiform, filamentos). De cele mai multe ori protalul se aseamănă cu talul algelor verzi filamentoase. Protalul se fixează de substrat prin intermediul rizoizilor de origine epidermică. Indiferent de formă şi mărime protalul se dezvoltă întotdeauna separat sau independent de sporofit, pe suporturi permanent umede. Pe protale se diferenţiază organele de reproducere, arhegoanele şi anteridiile la ferigile izospore sau arhegoane pe megaprotale şi anteridii pe microprotale în cazul ferigilor heterospore. Fecundaţia şi formarea zigotului. Ca la toate plantele cu gameţi masculini mobili, fecundaţia se produce în mediu lichid. Graţie flagelilor, spermatozoizii ajung la arhegoane, pătrund în interiorul lor şi fecundează oosfera. După fuzionarea celor doi gameţi haploizi se formează zigotul, germene sexuat diploid. Din zigot, în urma a numeroase diviziuni, se formează embrionul, iar din acesta o nouă plantă. Mult timp reproducerea sexuată a pteridofiţelor a rămas necunoscută, motiv, pentru care ele au fost denumite şi criptogame (vasculare). Ciclul de dezvoltare la pteridofite. Cele două generaţii, sporofitul şi gametofitul, îşi duc viaţa independent, alternând una cu cealaltă. Generaţia sporofitică dominantă este reprezentată de planta propriu-zisă. Pe aceasta se diferenţiază sporangii în care se formează sporii haploizi. Din germinarea sporilor rezultă gametofitul, numitprotal, pe care se diferenţiază gametangii masculini şi feminini, în urma fecundaţiei dintre gameţii de sex opus se formează zigotul iar din acesta embrionul (sporofitul tânăr) şi apoi sporofitul de lungă durată. Un astfel de ciclu de dezvoltare, în care alternează două generaţii morfologic diferite, independente, cu ploidii diferite, poartă numele de ciclu digenetic, diplo – haplo - fazic, iar plantele respective diplo - haplonte. Ciclul de viaţă la ferigi se desfăşoară diferit la cele izospore (Polypodium, Equisetum, Lycopodium), comparativ cu cele heterospore (Selaginella, Salvinia, Azolla) [4].
Aspectul exterior al
ferigilor. Cea mai
importantă adaptare apărută în morfologia ferigilor o
constituie diferenţierea corpului acestora în rădăcină, tulpină şi frunză,
toate alcătuind cormul. Diferenţierea morfologica a cormului s-a făcut în
paralel cu una structural/anatomică. Spre deosebire de talofite, la ferigi au apărut numeroase
ţesuturi şi s-a generalizat reproducerea sexuată. În aceste condiţii era nevoie
ca acest corp vegetativ imens comparativ cu talul să fie aprozivionat cu apă şi
hrănit. Astfel s-au diferenţiat ţesuturile conducătoare, specializate în
conducerea apei şi sărurilor minerale (ţesuturi conducătoare lemnoase) şi a
sevei elaborate (ţesuturi conducătoare liberiene). Ţesutul lemnos alcătuit
predominant din traheide cu punctuaţiuni scalariforme, este organizat în
fascicule conducătoare: un singur fascicul (Selaginella), două (Dryopteris,
Polypodium s.a.) sau mai multe (Pteridium, Lycopodium s.a.). Ţesutul liberian
alcătuit din celule ciuruite lipsite de celule anexe este organizat şi el în
fascicule conducătoare, în număr egal cu al celor lemnoase. Totalitatea
fasciculelor conducătoare formează cilindrul central (stei); la ferigi se
întâlnelşte în mod frecvent protostelul. Pteridofitele sunt primele cormofite
în a căror structură întâlnim ţesuturi conducătoare diferenţiate în elemente
lemnoase şi liberiene; din această cauză au mai fost denumite şi criptogame vasculare
sau traheofite. Asemănător altor plante care se reproduc sexuat, în viaţa
ferigilor se succed două generaţii: sporofitul (S) şi gametofitul (G). Sporofitul este generaţia diploidă
(2n) producătoare de spori. El îşi începe existenţa cu zigotul
(după fecundaţie) şi se încheie cu formarea sporilor haploizi, după meioză. La
ferigi, generaţia dominantă este sporofitul reprezentat de planta însăşi, alcătuită
din rădăcină, tulpină şi frunză. 1. Rădăcina îşi are originea în radicula
embrionului, întrucât radicula are o viaţă foarte scurtă, la ferigile mature nu
găsim, de obicei, rădăcină principală. Rădăcinile obişnuite sunt cele
adventive, care se formează pe rizomi. Spre deosebire de alte rizofite,
rădăcina unor ferigi (Lycopodium, Selaginella, Isoetes) se ramifică dihotomic.
Alte ferigi au rădăcini monopodiale.
2. Tulpina multor ferigi actuale este
reprezentată de un rizom de tip monopodial. Tulpinile supraterane pot fi
dihotomice (Lycopodium complanatum), simpodiale (Selaginella) sau monopodiale.
Adesea sunt asimilatoare, preluând rolul frunzelor (Eqttisetum sp.). Din punct
de vedere al structurii lor, ferigile ocupă un loc aparte. Cele mai multe
ferigi sunt perixile; în acest caz locul măduvei este ocupat de fasciculele
lemnoase unite prin bazele lor. Fasciculele liberiene se unesc la rândul lor,
în jurul lemnului, formând un manşon mai mult sau mai puţin complet (Polypodium
vulgare). Alte ferigi (Ophioglossum) au structură centroxilă (caracteristică
fanerogamelor). Un caracter important al tulpinii multor ferigi îl reprezintă
polistelia, care constă în prezenţa mai multor steluri, (meristeluri), fiecare
alcătuit din mai multe „fascicule conducătoare" delimitate de un periciclu
şi o endodermă proprie. Ferigile actuale nu posedă structură secundară [4].
3. Frunza ferigilor este foarte variabilă din
punct de vedere morfologic. Toate ferigile sunt autotrofe, funcţia de fotosinteză sau
asimilaţie clorofiliană, revenind frunzei. Din punct de vedere morfologic,
ferigile actuale posedă două tipuri de frunze, microfile şi macrofile. Primul
tip caracterizează ferigile microfilme, cu frunze mici scvamifonne sau
ligulate, cu excepţia lui Isoetes, care are frunze lungi şi subuliforme. La
majoritatea speciilor sunt dispuse atât de apropiat (spiralat) încât
internodurile nu se mai recunosc. Numai la Psilotum frunzele sunt mai distanţate.
La Equisetatae frunzele sunt dispuse verticilat în aşa fel încât atât nodurile
cât şi internodurile sunt vizibile. La Lycopodium, după desprinderea frunzelor,
rămân pe tulpini bazele acestora sub formă de mici proeminenţe; la Eqiiisetum,
frunzele dispuse în verticil, fuzionează prin bazele lor, formând o teacă
foliară. Ferigi microfilme întâlnim în clasele Lycopodiatae şi Equisetatae. Al
doilea tip caracterizează ferigile numite macrofiline din Polypodiatae.
Speciile din această clasă posedă frunze mari cu configuraţie diversificată,
numite şi fronde. O macrofilă este alcătuită, de obicei, din baza frunzei mai
dilatată, peţiol (stip) şi lamina, adesea divizată. Lamina prezintă aspecte
variate; ea poate fi întreagă (Pliyllitis) sau penat-compiisă. Frunzele penate
sunt alcătuite din ramificaţii foliate de ordinul I numite segmente primare,
lobi sau aripi (pinna), dispuse pe ambele laturi ale unei axe comune numită
rahis; ele pot, la rândul lor, să prezinte ramificaţii foliate de ordinul II
(segmente secundare); acestea se pot ramifica în segmente foliate de ordinul
III (segmente terţiare) şi chiar de ordinul IV. Elementele foliate ale
segmentelor de ordinul II-IV se numescpimde (aripioare). Uneori cu greu se
poate distinge lamina de peţiol, ceea ce este valabil şi pentru segmente.La
Pilularia frunza este reprezentată de un organ acicular care corespunde unui
peţiol fără lamina. Ramificarea segmentelor se face de obicei într-un plan şi
anume în două direcţii. La Ophioglossum s.a. segmentul foliar fertil este
altfel orientat decât cel steril. Din punct de vedere fiziologic, la unele
ferigi frunzele pot fi asimilatoare şi sporangifere (Ophioglossum, Botrichytim
s.a); primele se numesc trofofile, celelalte sporofile. Există însă şi o a
treia categorie de frunze trofosporofile; ele îndeplinesc ambele funcţii
(Dryopteris, Cystopteris, Phyllitis s.a). Din punct de vedere morfologic se
deosebesc frunze întregi (Phyllitis, Salvinia), penat-partite (Polypodium),
penat-sectate (Asplenium trichomanes), dublu-penat-sectate
(Athyriumfilix-femina s.a). Nervaţiunea este şi ea diferită: uninervă la
ferigile microfilme (Selaginella, Lycopodittm) sau multinervă la cele
macrofiline. Aceasta din urmă poate fi, penată (Polypodium vulgare) sau foarte
rar palmată (Pteridium) [4].
Evoluția.
Ferigile sunt
considerate primele cormofite apărute pe uscat, adaptate structural şi
funcţional mediului terestru şi mai rar celui acvatic. Pentru specialişti,
ferigile actuale descendente ale celor Paleozoice, de acum 350 milioane de ani,
reprezintă un grup de plante din care au derivat plantele cu seminţe. Cei mai
mulţi specialişti consideră că ferigile îşi au originea în algele verzi. Fergile au avut o dezvoltare luxuriantă în Paleozoicul
mediu şi superior, denumit din acest motiv şi era pteridofitelor. Unele dintre aceste plante vechi creşteau la înălţimi
mari, şi chiar dacă ferigile arborescente înalte de 30 m, şi pedicuţele înalte
de 15 m au dispărut, mai există în prezent varietăţi mai mici. Alături de
specii evoluate mai recent.
În vremurile modeme, ferigile sunt utilizate
de oameni doar ca plante decorative, dar plantele masive care au existat cu
milioane de ani în urmă au cauzat formarea cărbunelui şi ţiţeiului, ambele
resurse esenţiale pentru creşterea şi dezvoltarea erei moderne.
Ferigile,
pedicuţele şi coada calului fac parte din grupul cel mai avansat de plante fără
flori. Spre deosebire de muşchii frunzoşi şi muşchii pieloşi, care au rizoizi,
sau peri. Servind drept rădăcini, ferigile şi plantele înrudite cu ele au rădăcini
adevărate cu un sistem funcţional de vase, sau tuburi, pentru transportul apei,
şi frunze cu o structură la fel de avansată ca şi a multora dintre plantele cu
flori [2].
Este necesar
de menționat că ferigile sunt adevărate fosile vii. Deși există și varietăți
apărute în epocile geologice mai
recente, forma și structura morfologică la majoritatea nu s-a schimbat
semnificativ sau chiar de loc. Pentru ferigi lumea s-a oprit acum 180 de
milioane de ani.
Se spune că atunci cînd un organism nu își modifică
structura timp de milioane de ani, se poate ajunge la concluzia că natura a
calculat la perfectie genotipul și i-a dat… cea mai bună formă posibilă. O
recentă descoperire facută în Suedia a arătat că ferigile și-au încetat
evoluția acum 180 de milioane de ani.
S-a ajuns
la aceasta constatare după ce o astfel de plantă, cu o lungime de 5.8 cm si
lata de 4.1 cm, a fost scoasă la lumină dintr-un depozit mineralier perfect
fosilizată. Organismul este atît de bine conservat încît se pot observa cu
ușurință nu numai frunzele, ci și celulele, nucleele celulare sau legăturile
dintre ele. Analiza la nivel subcelular a arătat o similaritate cu ferigile
actuale în ce privește mărimea nucleelor, conținutul ADN și numărul
cromozomilor.

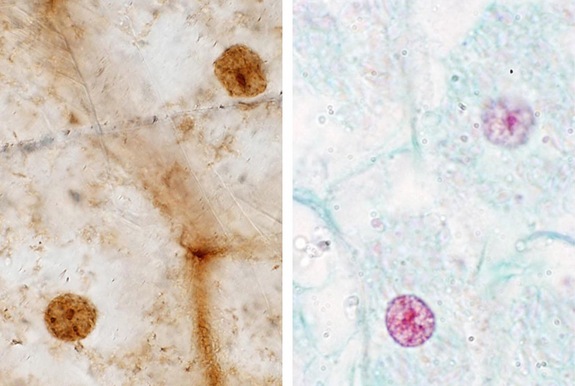
Figura nr. 2. Structura subcelulară la frunzele
ferigilor, imaginea din stânga este amprenta pietrificată a frunzei de ferigă,
iar în partea dreaptă este structura subcelulară a ferigilor contemporane [2].
Așadar
dacă vă veți întreba vreodată cum arăta o pădure de acum 180 de milioane de
ani, imaginați-vă o pădure de la cota 1500. Aproape că nu s-a schimbat nimic [2].
Importanța și rolul ferigilor pentru om. În ultimul deceniu ferigile au
intrat în atenţia marelui public, în special a iubitorilor de plante
ornamentale din cel puţin două motive: - având în vedere sempervirescenţa lor,
un număr din ce în ce mai mare de specii de ferigi este folosit ca plante
decorative, în special de apartament, dar şi de grădină. Aşa este cazul
ferigilor: Dryopteris, Cystopteris, Polypodium, Selaginella, Platyceritim,
Asplenium ş.a. - numeroase specii au fost şi sunt utilizate ca plante medicinale.
Dintre acestea sunt: Equisetum arvense (ca diuretic, hemostatic, remineralizant,
antidiareic), Polypodium vulgare (ca antihelmintic, colagog, laxativ,
expectorant ş.a.), Dryopteris felix femina (antihelmintic), Lycopodium clavatum
(hemostatic) s.a. O atenţie deosebită se acordă, în zilele noastre, protecţiei
şi conservării ferigilor, cunoaşterii biologiei acestora şi stopării degradării
populaţiilor de ferigi. Defrişarea pădurilor, locul de viaţă al ferigilor,
vânzarea frunzelor de ferigi în buchete de flori tăiate afecteză permanent
speciile de ferigi din flora spontană.
Bibliografie:
la 31.05.2014).
5. 3.bp.blogspot.com/-ewxTpakLx9I/TfB2wy03wNI/AAAAAAAABWs/nNGL2u4SRLk/s1600/IMG_0289.JPG (accesat la 23.09.2015).
Abonați-vă la:
Comentarii (Atom)